Dluzewska J, Dziegielewski W, Szymanska-Lejman M, Gazecka M, Henderson IR, Higgins JD, Ziolkowski PA. MSH2 stimulates interfering and inhibits non-interfering crossovers in response to genetic polymorphism. Nat Commun 14, 6716 (2023)
Mejoza to podział komórkowy niezbędny do rozmnażania płciowego organizmów. W jej wyniku dochodzi do redukcji liczby chromosomów w powstających gametach, a pełny zestaw informacji genetycznej zostaje odtworzony dopiero po zapłodnieniu. Co ważne, zanim liczba chromosomów zostanie zredukowana dochodzi do przetasowania materiału genetycznego w procesie rekombinacji crossing-over, który polega na wzajemnej wymianie fragmentów chromosomów homologicznych pochodzących od matki i ojca. Możliwość kontroli crossing-over jest bardzo ważna dla hodowców, ponieważ umożliwia transfer genów kodujących pożądane cechy pomiędzy różnymi odmianami.
Jednym z czynników wpływających na decyzję o formowaniu crossing-over jest obecność polimorfizmów, czyli różnic w DNA pomiędzy rekombinującymi chromosomami, które są rozpoznawane przez kompleksy MSH2. W komórkach somatycznych białko MSH2 posiada właściwości antyrekombinacyjne. Natomiast rola MSH2 w mejozie nie jest do końca poznana.
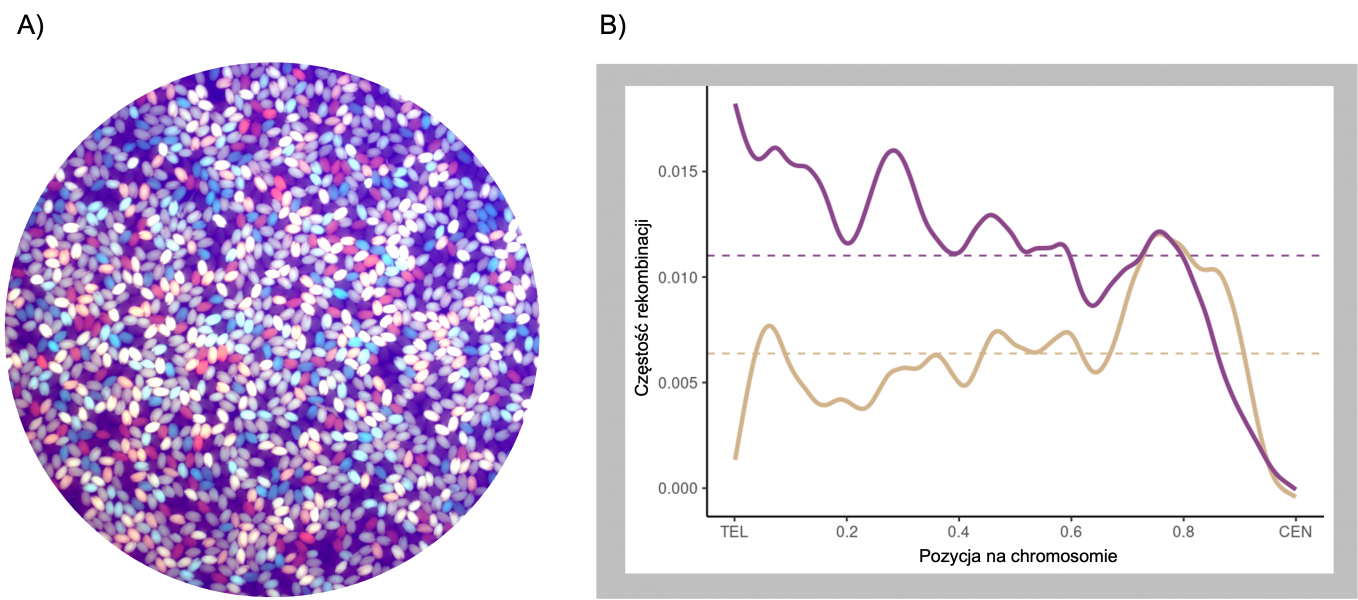
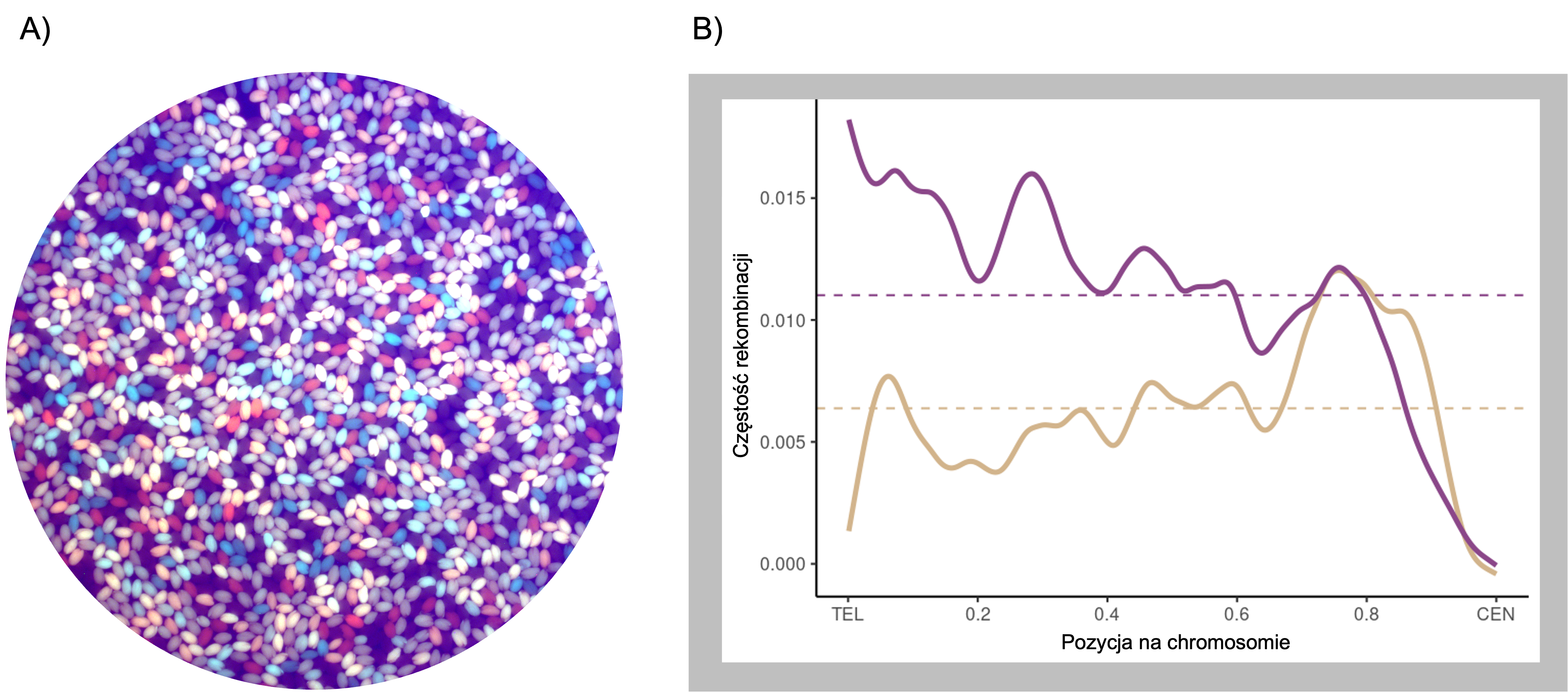
Zdarzenia crossing-over są formowane w komórce na drodze dwóch niezależnych szlaków, w związku z czym dzieli się je na crossing-over klasy I oraz klasy II. Poprzez wyłączenie poszczególnych szlaków zbadaliśmy częstość rekombinacji oddzielnie w kontekście klasy I oraz klasy II, zarówno w konkretnych regionach chromosomowych w oparciu o system fluorescencyjnych nasion (Rycina A), jak i w skali całego genomu (Rycina B). W ten sposób odkryliśmy, iż w roślinie modelowej Arabidopsis thaliana, białko MSH2, w odpowiedzi na obecność polimorfizmów pomiędzy rekombinującymi chromosomami, stymuluje formowanie crossing-over klasy I, ale blokuje klasę II. Antyrekombinacyjna rola MSH2 w klasie II w regionach polimorficznych polega prawdopodobnie na stymulacji działania helikaz DNA, które powodują rozpad produktów pośrednich rekombinacji i przez to naprawę przy pomocy innych mechanizmów. Ten antagonistyczny wpływ MSH2 na oba szlaki crossing-over prawdopodobnie wiąże się z ich różną funkcją biologiczną: Klasa I jest specyficzna dla mejozy i odpowiada za większość powstających crossing-over. W mejozie stymulacja crossing-over w regionach polimorficznych umożliwia przetasowanie informacji genetycznej, a w konsekwencji powstanie nowych genotypów, co jest ważne w adaptacji i ewolucji genomów. Natomiast białka odpowiedzialne za powstawanie crossing-over klasy II działają głównie w komórkach somatycznych, gdzie ich funkcją jest naprawa DNA. Dlatego klasa II nie jest aktywna w regionach polimorficznych, gdzie mogłaby prowadzić do mutacji.
Co więcej, przeanalizowaliśmy, jak rozkład polimorfizmu wzdłuż chromosomu wpływa na rekombinację. Okazuje się, że wzór rekombinacji jest podobny w liniach wsobnych, które są pozbawione polimorfizmów oraz w mieszańcach, które posiadają setki tysięcy polimorfizmów wzdłuż chromosomów. Natomiast wzór crossing-over zmienia się dramatycznie, gdy fragment chromosomu jest polimorficzny podczas gdy reszta jest niepolimorficzna – wtedy dominujące crossing-over klasy I redystrybuowane są do regionów z polimorfizmami. Dlaczego rozkład rekombinacji jest podobny w liniach wsobnych i mieszańcach? Otóż u osobników wsobnych, gdy nie ma polimorfizmów, MSH2 nie ma do czego się wiązać i rozkład crossing-over zależy głównie od położenia na chromosomie. Z kolei u mieszańców polimorfizmy występują na całej długości chromosomu, kompleksy MSH2 wiążą się do nich dość równomiernie i w konsekwencji znów głównie położenie na chromosomie wpływa na rozmieszczenie crossing-over klasy I. Jednakże w sytuacji, gdy pojedynczy region polimorficzny zlokalizowany jest na niepolimorficznym chromosomie, dochodzi do lokalnego wzrostu stężenia kompleksów MSH2 rozpoznających niedopasowania i stymulację crossing-over klasy I w regionie polimorficznym.